您可以通过点击 CST 信号转导通路图的各个节点查找研究资源或产品信息。您还可以下载通路图,将其用于教育和研究。
染色质调控是指影响染色质结构的活动,因此也影响基因表达模式的翻译调控。
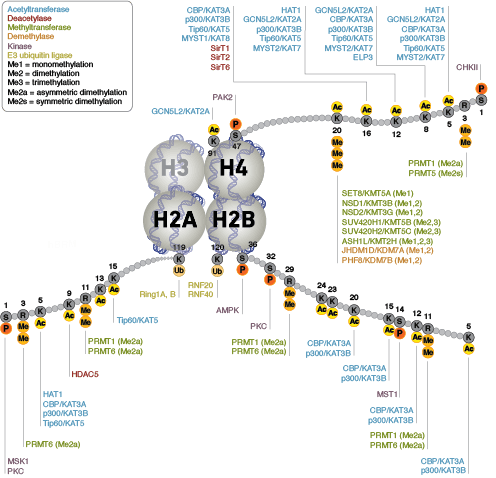
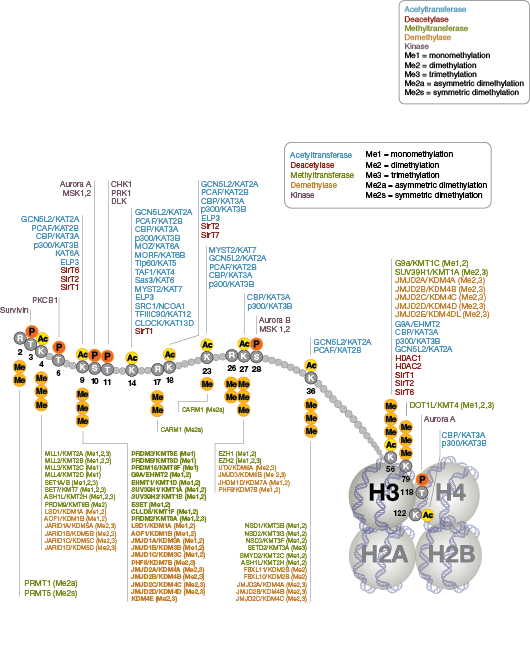
表观遗传学特指会造成基因表达发生改变且不在 DNA 内进行编码的遗传修饰。核小体,由四个组蛋白组成(H2A、H2B、H3 和 H4),是染色质的主体结构。最初认为组蛋白具有用作 DNA 包装的静态支架的功能,但最近已证明,组蛋白是动态的蛋白质,它经历了多种类型的翻译后修饰 (PTMs),并与调节蛋白相互影响来控制基因表达。
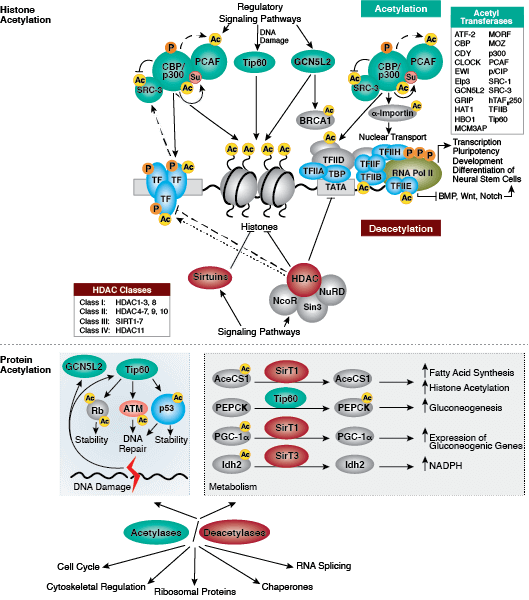
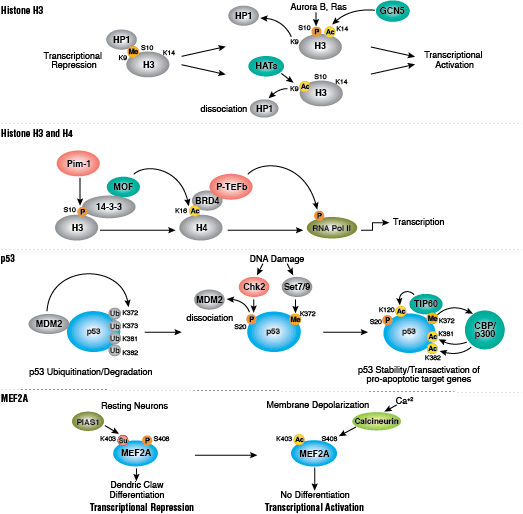
蛋白质乙酰化在调节染色质结构和转录活动中发挥关键作用。组蛋白乙酰转移酶 (HAT) 作用下的组蛋白高度乙酰化与转录激活相关,而组蛋白去乙酰化酶 (HDAC) 作用下的组蛋白去乙酰化则与转录抑制相关。高度乙酰化可在中和组蛋白尾部的正电荷以及扰乱核小体之间、核小体与 DNA 之间的相互作用的情况下,直接影响染色质结构。此外,乙酰化会为含有 bromodomain 的染色质调节蛋白(蛋白修饰阅读器)创造结合位点。
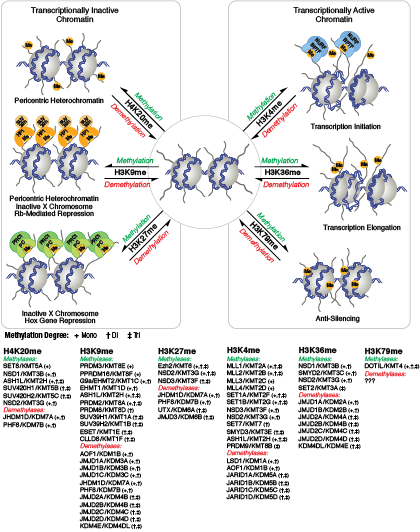
与乙酰化不同的是,甲基化不会改变精氨酸和赖氨酸残基的电荷,也不可能直接调节染色质折叠所需的核小体之间的相互作用。甲基化的精氨酸与赖氨酸残基是形成基因组的活性区与非活性区的主要决定因素。甲基化可促进染色质调节蛋白/组蛋白修饰阅读器的结合,其中包含多种甲基赖氨酸或甲基精氨酸结合域(PHD 域、chromo 域、WD40 域、Tudor 域、MBT 域、锚蛋白重复序列域、PWWP 域)。辅助活化蛋白和辅助抑制蛋白的募集依赖于被修饰的特定赖氨酸残基。
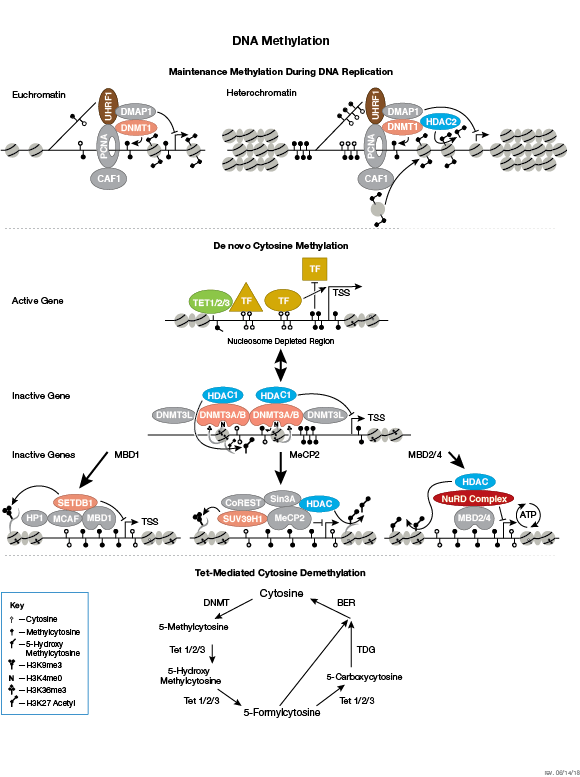
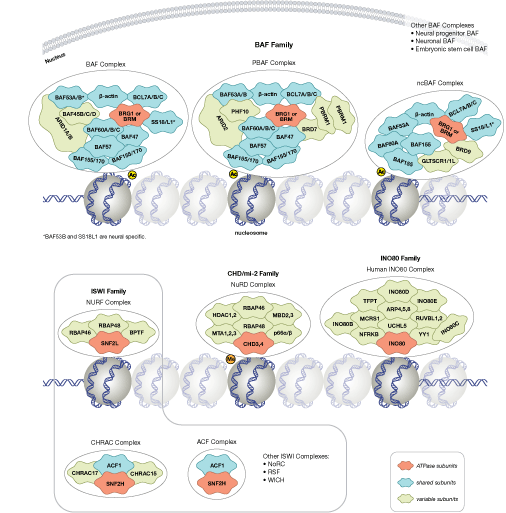
调节染色质结构是调控转录激活和抑制的必要部分。一个可以调节染色质结构的办法是,通过 ATP 依赖的染色质重塑物(例如:NuRD、Polycomb 和 SWI/SNF 复合体)破坏组蛋白-DNA 的接触。这些重塑物已被证实能够调控基因活化/抑制、细胞生长、细胞周期和分化。染色质结构也能够被其他 PTM 调控,如组蛋白的磷酸化,它能够影响与 DNA 相互作用的蛋白的关联,并在最近已被证实在协调其他组蛋白修饰中发挥作用。另外,哺乳动物细胞中胞嘧啶残基上的 DNA 甲基化能够影响染色质折叠,是一种可遗传的表观遗传修饰,该修饰对于基因沉默、基因组印记和发育的正确调控都是至关重要的。三大哺乳动物的 DNA 甲基转移酶家族已被识别出来,即 DNMT1/2/3,在胚胎干细胞和成年体细胞中发挥各自的作用。除了核心组蛋白,还存在许多组蛋白变体。它们赋予核小体不同的结构性质,并发挥许多特定功能,例如:DNA 修复、在有丝分裂期间正确组装着丝粒和染色体分离以及转录调控。
染色质和表观遗传调节对于在发育期间和处于压力条件下的基因组正确编码至关重要,因为基因表达的误调节会导致疾病状态,如癌症。