凋亡、线粒体自噬、坏死性凋亡和自噬等细胞死亡通路发生突变都会导致神经元细胞死亡及神经退行性疾病进展。促凋亡和抗凋亡信号转导异常、线粒体功能障碍、自噬误调节或未折叠蛋白反应以及在应激和/或炎症刺激下的坏死体激活都只强调了神经元死亡或变得有病变的少数几种机制。虽然人们了解非神经元细胞中的许多这些通路,但其在有自身问题的神经元中的激活和失调机制仍然是一个谜。
在氧化应激的刺激下,PARP 通常在 DNA 修复通路中起到关键作用。在 Asp214 和 Gly215 之间被 caspase 3 裂解活化时,氨基末端裂解片段会抑制 DNA 修复酶将神经元推向凋亡过程,从而使其成为凋亡细胞的一个标志。
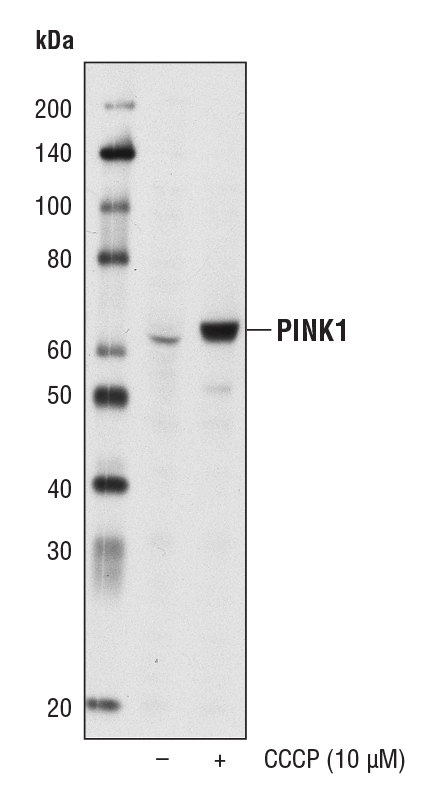
PTEN 诱导性激酶 1 (PINK1) 是一种线粒体丝氨酸/苏氨酸蛋白激酶,可避免细胞出现应激诱导的线粒体功能障碍。它聚集在严重受损线粒体的外膜上,并募集 PARKIN 以通过自噬诱导降解。PINK1 突变与常染色体隐性早发型帕金森病有关。
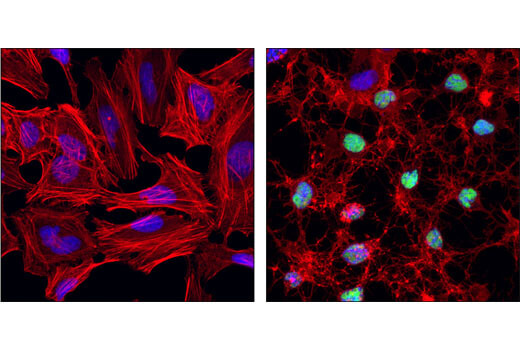
Sequestosome 1 (SQSTM1, p62) 是一种自噬体转运蛋白,可结合蛋白聚合物以靶向它们进行选择性自噬。SQSTM1/p62 突变会导致 α-突触核蛋白、亨廷顿蛋白、Tau 蛋白以及 β-淀粉样蛋白在细胞内的聚集增加,从而分别促成帕金森病、亨廷顿氏舞蹈病和阿尔茨海默病的进展。
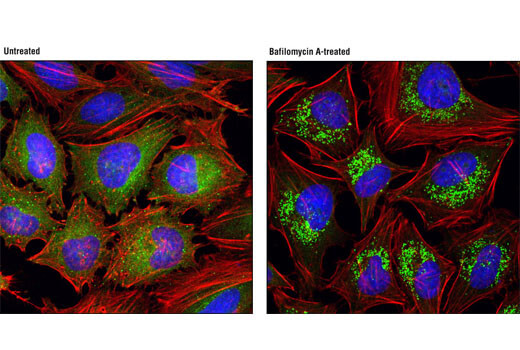
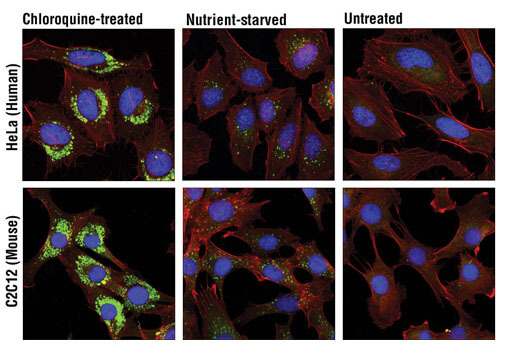
LC3A/B 在自噬体生物发生和成熟中起到关键作用,并且还作为一种接头蛋白选择性募集转运蛋白到自噬体上。在发生 TREM2 突变的阿尔茨海默病患者组织中观察到 LC3 阳性小胶质细胞增多,表明 TREM2 依赖性自噬中断会促成阿尔茨海默病的病因。
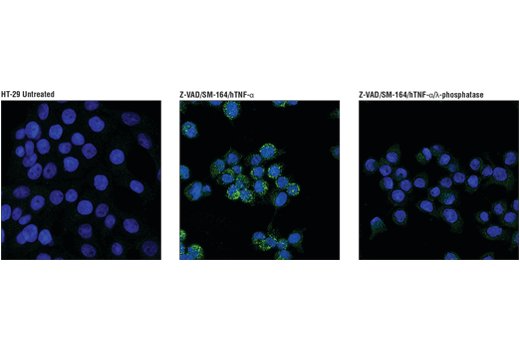
Human Phospho-RIP3 (Ser227) 使 MLKL1 磷酸化,从而触发 TNF 诱导的坏死性凋亡。这种形式的程序性细胞死亡已在多发性硬化和肌萎缩性脊髓侧索硬化症中报道。