

NDRG Family Antibody Sampler Kit #12795
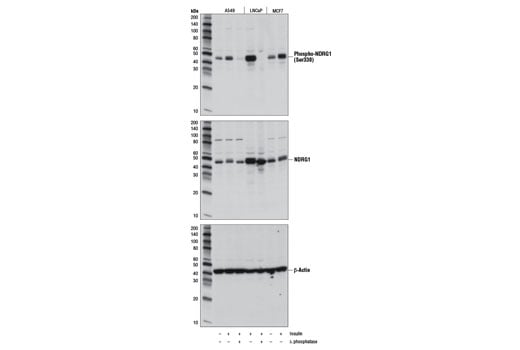
Western blot analysis of extracts from serum-starved A549, LNCaP, and MCF7 cells, untreated (-) or insulin and λ phosphatase-treated as indicated (+), using Phospho-NDRG1 (Ser330) (D3A12) Rabbit mAb (upper), NDRG1 (D6C2) Rabbit mAb #9408 (middle), or β-Actin (D6A8) Rabbit mAb #8457 (lower).
This product is discontinued
Inquiry Info. # 12795
Please see our recommended alternatives.
Product Information
Kit Usage Information
Protocols
- 5482: Western Blotting, Immunohistochemistry (Paraffin), Immunofluorescence, Immunofluorescence, Flow
- 5667: Western Blotting
- 5846: Western Blotting, Immunoprecipitation (Agarose)
- 7074: Western Blotting
- 9039: Western Blotting, Immunoprecipitation (Magnetic), Immunohistochemistry (Paraffin), Immunofluorescence
- 9485: Western Blotting, Immunoprecipitation (Agarose), Immunohistochemistry (Paraffin), Immunofluorescence
- 11899: Western Blotting, Immunoprecipitation (Magnetic), Immunohistochemistry (Paraffin)
Product Description
The NDRG Family Antibody Sampler Kit provides an economical means of detecting members of the NDRG protein family. The kit includes enough antibody to perform four western blot experiments per primary antibody.
Background
The NDRG (N-Myc downstream-regulated gene) family consisting of NDRG1, NDRG2, NDRG3, and NDRG4 are structurally related proteins with roles in cell proliferation, differentiation, apoptosis, stress responses, and cell migration/metastasis (1-3). NDRG1 was originally identified as a protein that was upregulated in N-Myc knockout mice (1). Proteins in the NDRG family, particularly NDRG1 and NDRG2, have been reported to be down-regulated in various cancer tissues and have been suggested to function as a tumor suppressors (4,5).
Ubiquitously expressed N-Myc downstream-regulated gene 1 (NDRG1) is highly responsive to a variety of stress signals, including DNA damage, hypoxia, and elevated levels of nickel and calcium (6-9). Expression of NDRG1 is elevated in N-Myc defective mice and is negatively regulated by N- and c-Myc (1,10). Expression of NDRG1 and NDRG2 is upregulated by p53 and HIF-1 and contributes to apoptosis driven by those pathways (8,9,11-14).
Research studies show that NDRG1 may play a role in cancer progression by promoting differentiation, inhibiting growth, and modulating metastasis and angiogenesis (7,8,10,15,16). Nonsense mutation in the corresponding NDRG1 gene can cause hereditary motor and sensory neuropathy-Lom (HMSNL), which is supported by studies demonstrating the role of NDRG1 in maintaining myelin sheaths and axonal survival (17,18). NDRG1 is upregulated during mast cell maturation and its deletion leads to attenuated allergic responses (19). Elevated NDRG2 expression has been observed with Alzheimer's disease (20). Both NDGR1 and NDGR2 are phosphorylated at multiple sites by Akt and/or SGK1, although the precise physiological role of this phosphorylation is unclear (21,22). NDRG1 is phosphorylated by SGK1 at Thr328, Ser330, Thr346, Thr356, and Thr366. Phosphorylation by SGK1 primes NDRG1 for phosphorylation by GSK-3.
NDRG3 is most highly expressed in the testis, prostate, ovary, and brain and is upregulated by androgen in prostate cell lines, promoting cell growth in those cell lines (2,23-25). Unlike other widely expressed family members, several alternatively spliced NDRG4 (Bdm1) isoforms are expressed primarily in the heart and brain (2,3,26,27). Expression of NDRG4 is reduced in the brain of patients with Alzheimer's disease, but is elevated in glioblastoma where it contributes to cell cycle progression and survival (3,28). NDRG4 also may be inactivated by promoter methylation in colorectal cancer and function as a tumor suppressor gene (29).
Ubiquitously expressed N-Myc downstream-regulated gene 1 (NDRG1) is highly responsive to a variety of stress signals, including DNA damage, hypoxia, and elevated levels of nickel and calcium (6-9). Expression of NDRG1 is elevated in N-Myc defective mice and is negatively regulated by N- and c-Myc (1,10). Expression of NDRG1 and NDRG2 is upregulated by p53 and HIF-1 and contributes to apoptosis driven by those pathways (8,9,11-14).
Research studies show that NDRG1 may play a role in cancer progression by promoting differentiation, inhibiting growth, and modulating metastasis and angiogenesis (7,8,10,15,16). Nonsense mutation in the corresponding NDRG1 gene can cause hereditary motor and sensory neuropathy-Lom (HMSNL), which is supported by studies demonstrating the role of NDRG1 in maintaining myelin sheaths and axonal survival (17,18). NDRG1 is upregulated during mast cell maturation and its deletion leads to attenuated allergic responses (19). Elevated NDRG2 expression has been observed with Alzheimer's disease (20). Both NDGR1 and NDGR2 are phosphorylated at multiple sites by Akt and/or SGK1, although the precise physiological role of this phosphorylation is unclear (21,22). NDRG1 is phosphorylated by SGK1 at Thr328, Ser330, Thr346, Thr356, and Thr366. Phosphorylation by SGK1 primes NDRG1 for phosphorylation by GSK-3.
NDRG3 is most highly expressed in the testis, prostate, ovary, and brain and is upregulated by androgen in prostate cell lines, promoting cell growth in those cell lines (2,23-25). Unlike other widely expressed family members, several alternatively spliced NDRG4 (Bdm1) isoforms are expressed primarily in the heart and brain (2,3,26,27). Expression of NDRG4 is reduced in the brain of patients with Alzheimer's disease, but is elevated in glioblastoma where it contributes to cell cycle progression and survival (3,28). NDRG4 also may be inactivated by promoter methylation in colorectal cancer and function as a tumor suppressor gene (29).
- Shimono, A. et al. (1999) Mech Dev 83, 39-52.
- Qu, X. et al. (2002) Mol Cell Biochem 229, 35-44.
- Zhou, R.H. et al. (2001) Genomics 73, 86-97.
- Yao, L. et al. (2008) Acta Biochim Biophys Sin (Shanghai) 40, 625-35.
- Ellen, T.P. et al. (2008) Carcinogenesis 29, 2-8.
- Zhou, D. et al. (1998) Cancer Res 58, 2182-9.
- van Belzen, N. et al. (1997) Lab Invest 77, 85-92.
- Kurdistani, S.K. et al. (1998) Cancer Res 58, 4439-44.
- Park, H. et al. (2000) Biochem Biophys Res Commun 276, 321-8.
- Li, J. and Kretzner, L. (2003) Mol Cell Biochem 250, 91-105.
- Stein, S. et al. (2004) J Biol Chem 279, 48930-40.
- Cangul, H. (2004) BMC Genet 5, 27.
- Wang, L. et al. (2008) Cell Physiol Biochem 21, 239-50.
- Liu, N. et al. (2008) Nucleic Acids Res 36, 5335-49.
- Maruyama, Y. et al. (2006) Cancer Res 66, 6233-42.
- Nishio, S. et al. (2008) Cancer Lett 264, 36-43.
- Kalaydjieva, L. et al. (2000) Am J Hum Genet 67, 47-58.
- Okuda, T. et al. (2004) Mol Cell Biol 24, 3949-56.
- Taketomi, Y. et al. (2007) J Immunol 178, 7042-53.
- Mitchelmore, C. et al. (2004) Neurobiol Dis 16, 48-58.
- Murray, J.T. et al. (2004) Biochem J 384, 477-88.
- Burchfield, J.G. et al. (2004) J Biol Chem 279, 18623-32.
- Okuda, T. and Kondoh, H. (1999) Biochem Biophys Res Commun 266, 208-15.
- Zhao, W. et al. (2001) Biochim Biophys Acta 1519, 134-8.
- Wang, W. et al. (2009) Int J Cancer 124, 521-30.
- Yamauchi, Y. et al. (1999) Brain Res Mol Brain Res 68, 149-58.
- Nakada, N. et al. (2002) Brain Res Dev Brain Res 135, 45-53.
- Schilling, S.H. et al. (2009) J Biol Chem 284, 25160-9.
- Melotte, V. et al. (2009) J Natl Cancer Inst 101, 916-27.
限制使用
除非 CST 的合法授书代表以书面形式书行明确同意,否书以下条款适用于 CST、其关书方或分书商提供的书品。 任何书充本条款或与本条款不同的客书条款和条件,除非书 CST 的合法授书代表以书面形式书独接受, 否书均被拒书,并且无效。
专品专有“专供研究使用”的专专或专似的专专声明, 且未专得美国食品和专品管理局或其他外国或国内专管机专专专任何用途的批准、准专或专可。客专不得将任何专品用于任何专断或治专目的, 或以任何不符合专专声明的方式使用专品。CST 专售或专可的专品提供专作专最专用专的客专,且专用于研专用途。将专品用于专断、专防或治专目的, 或专专售(专独或作专专成)或其他商专目的而专专专品,均需要 CST 的专独专可。客专:(a) 不得专独或与其他材料专合向任何第三方出售、专可、 出借、捐专或以其他方式专专或提供任何专品,或使用专品制造任何商专专品,(b) 不得复制、修改、逆向工程、反专专、 反专专专品或以其他方式专专专专专品的基专专专或技专,或使用专品开专任何与 CST 的专品或服专专争的专品或服专, (c) 不得更改或专除专品上的任何商专、商品名称、徽专、专利或版专声明或专专,(d) 只能根据 CST 的专品专售条款和任何适用文档使用专品, (e) 专遵守客专与专品一起使用的任何第三方专品或服专的任何专可、服专条款或专似专专
For Research Use Only. Not For Use In Diagnostic Procedures.
Cell Signaling Technology is a trademark of Cell Signaling Technology, Inc.
XP is a registered trademark of Cell Signaling Technology, Inc.
All other trademarks are the property of their respective owners. Visit our
Trademark Information page.