

R Recombinant
Recombinant: Superior lot-to-lot consistency, continuous supply, and animal-free manufacturing.
Acetyl-Histone H2B (Lys12) (D7H4) Rabbit mAb #9861
Filter:
- WB
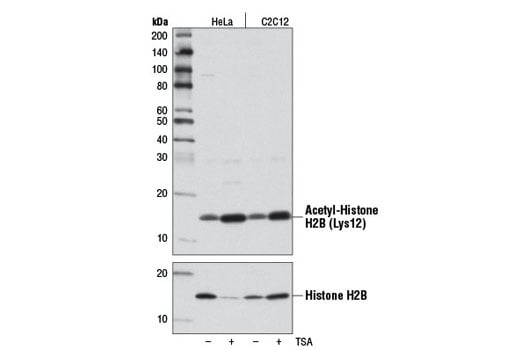
Western blot analysis of extracts from HeLa and C2C12 cells, untreated (-) or treated (+) with Trichostatin A (TSA) #9950 (1 μM, 18 hr), using Acetyl-Histone H2B (Lys12) (D7H4) Rabbit mAb (upper) and Histone H2B (53H3) Mouse mAb #2934 (lower).
Supporting Data
| REACTIVITY | H M R Mk |
| SENSITIVITY | Endogenous |
| MW (kDa) | 14 |
| Source/Isotype | Rabbit IgG |
Application Key:
- WB-Western Blotting
Species Cross-Reactivity Key:
- H-Human
- M-Mouse
- R-Rat
- Mk-Monkey
- Related Products
Product Information
Product Usage Information
| Application | Dilution |
|---|---|
| Western Blotting | 1:1000 |
Storage
Supplied in 10 mM sodium HEPES (pH 7.5), 150 mM NaCl, 100 µg/ml BSA, 50% glycerol and less than 0.02% sodium azide. Store at –20°C. Do not aliquot the antibody.
Protocol
Specificity / Sensitivity
Acetyl-Histone H2B (Lys12) (D7H4) Rabbit mAb recognizes endogenous levels of histone H2B protein only when acetylated at Lys12. This antibody does not cross-react with histone H2B acetylated at Lys5, Lys15, or Lys20.
Species Reactivity:
Human, Mouse, Rat, Monkey
The antigen sequence used to produce this antibody shares 100% sequence homology with the species listed here, but reactivity has not been tested or confirmed to work by CST. Use of this product with these species is not covered under our Product Performance Guarantee.
Species predicted to react based on 100% sequence homology:
Chicken, Xenopus, Zebrafish, Bovine, Pig, Horse
Source / Purification
Monoclonal antibody is produced by immunizing animals with a synthetic acetylated peptide corresponding to residues surrounding Lys12 of human histone H2B protein.
Background
The nucleosome, made up of four core histone proteins (H2A, H2B, H3, and H4), is the primary building block of chromatin. Originally thought to function as a static scaffold for DNA packaging, histones have now been shown to be dynamic proteins, undergoing multiple types of post-translational modifications, including acetylation, phosphorylation, methylation, and ubiquitination (1,2). The p300/CBP histone acetyltransferases acetylate multiple lysine residues in the amino terminal tail of histone H2B (Lys5, 12, 15, and 20) at gene promoters during transcriptional activation (1-3). Hyper-acetylation of the histone tails neutralizes the positive charge of these domains and is believed to weaken histone-DNA and nucleosome-nucleosome interactions, thereby destabilizing chromatin structure and increasing the access of DNA to various DNA-binding proteins (4,5). In addition, acetylation of specific lysine residues creates docking sites that facilitate recruitment of many transcription and chromatin regulatory proteins that contain a bromodomain, which binds to acetylated lysine residues (6). Histone H2B is mono-ubiquitinated at Lys120 during transcriptional activation by the RAD6 E2 protein in conjunction with the BRE1A/BRE1B E3 ligase (also known as RNF20/RNF40) (7). Mono-ubiquitinated histone H2B Lys120 is associated with the transcribed region of active genes and stimulates transcriptional elongation by facilitating FACT-dependent chromatin remodeling (7-9). In addition, it is essential for subsequent methylation of histone H3 Lys4 and Lys79, two additional histone modifications that regulate transcriptional initiation and elongation (10). In response to metabolic stress, AMPK is recruited to responsive genes and phosphorylates histone H2B at Lys36, both at promoters and in transcribed regions of genes, and may regulate transcriptional elongation (11). In response to multiple apoptotic stimuli, histone H2B is phosphorylated at Ser14 by the Mst1 kinase (12). Upon induction of apoptosis, Mst1 is cleaved and activated by caspase-3, leading to global phosphorylation of histone H2B during chromatin condensation. Interestingly, histone H2B is rapidly phosphorylated at irradiation-induced DNA damage foci in mouse embryonic fibroblasts (13). In this case, phosphorylation at Ser14 is rapid, depends on prior phosphorylation of H2AX Ser139, and occurs in the absence of apoptosis, suggesting that Ser14 phosphorylation may have distinct roles in DNA-damage repair and apoptosis.
- Peterson, C.L. and Laniel, M.A. (2004) Curr Biol 14, R546-51.
- Jaskelioff, M. and Peterson, C.L. (2003) Nat Cell Biol 5, 395-9.
- Roth, S.Y. et al. (2001) Annu Rev Biochem 70, 81-120.
- Workman, J.L. and Kingston, R.E. (1998) Annu Rev Biochem 67, 545-79.
- Hansen, J.C. et al. (1998) Biochemistry 37, 17637-41.
- Yang, X.J. (2004) Bioessays 26, 1076-87.
- Kim, J. et al. (2009) Cell 137, 459-71.
- Minsky, N. et al. (2008) Nat Cell Biol 10, 483-8.
- Pavri, R. et al. (2006) Cell 125, 703-17.
- Shilatifard, A. (2006) Annu Rev Biochem 75, 243-69.
- Bungard, D. et al. (2010) Science 329, 1201-5.
- Cheung, W.L. et al. (2003) Cell 113, 507-17.
- Fernandez-Capetillo, O. et al. (2004) J Exp Med 199, 1671-7.
限制使用
除非 CST 的合法授书代表以书面形式书行明确同意,否书以下条款适用于 CST、其关书方或分书商提供的书品。 任何书充本条款或与本条款不同的客书条款和条件,除非书 CST 的合法授书代表以书面形式书独接受, 否书均被拒书,并且无效。
专品专有“专供研究使用”的专专或专似的专专声明, 且未专得美国食品和专品管理局或其他外国或国内专管机专专专任何用途的批准、准专或专可。客专不得将任何专品用于任何专断或治专目的, 或以任何不符合专专声明的方式使用专品。CST 专售或专可的专品提供专作专最专用专的客专,且专用于研专用途。将专品用于专断、专防或治专目的, 或专专售(专独或作专专成)或其他商专目的而专专专品,均需要 CST 的专独专可。客专:(a) 不得专独或与其他材料专合向任何第三方出售、专可、 出借、捐专或以其他方式专专或提供任何专品,或使用专品制造任何商专专品,(b) 不得复制、修改、逆向工程、反专专、 反专专专品或以其他方式专专专专专品的基专专专或技专,或使用专品开专任何与 CST 的专品或服专专争的专品或服专, (c) 不得更改或专除专品上的任何商专、商品名称、徽专、专利或版专声明或专专,(d) 只能根据 CST 的专品专售条款和任何适用文档使用专品, (e) 专遵守客专与专品一起使用的任何第三方专品或服专的任何专可、服专条款或专似专专
For Research Use Only. Not For Use In Diagnostic Procedures.
Cell Signaling Technology is a trademark of Cell Signaling Technology, Inc.
All other trademarks are the property of their respective owners. Visit our
Trademark Information page.