您可以通过点击 CST 信号转导通路图的各个节点查找研究资源或产品信息。您还可以下载通路图,将其用于教育和研究。
核受体超家族是配体激活的转录因子,在细胞分化/发育、增殖和代谢中发挥不同作用,并且与多种病症有关,如癌症、心血管疾病、炎症及生殖异常。该家族的成员包含一个氨基末端反式激活结构域、一个高度保守的中央区域锌指 DNA 结合域以及一个 C端配体结合域。配体结合其相关的核受体会导致靶组织内特异性基因的反式激活。
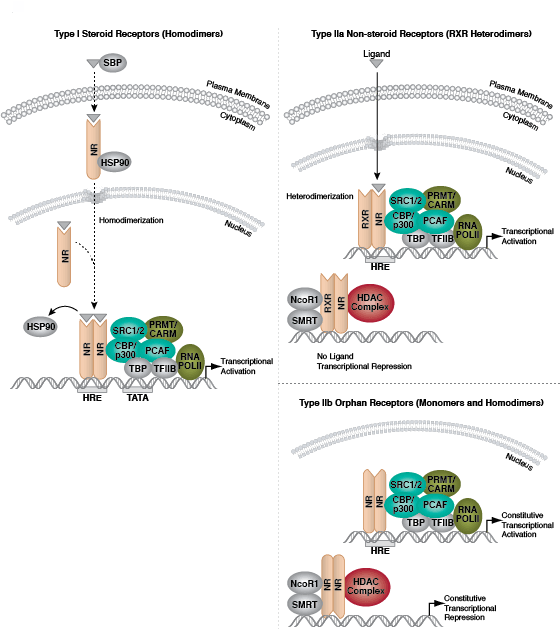
I 型核受体(也称类固醇受体)包括雌激素受体、雄激素受体、孕酮受体和糖皮质激素受体。这个受体亚组的类固醇激素配体从各自的内分泌腺通过血流转运后与类固醇结合球蛋白结合。在某种程度上,在细胞浆区室内结合各自的配体时,一些 I 型核受体被激活。配体-受体复合体进入胞核,在胞核中,复合体发生同源二聚化,从 HSP90 上分离,并结合某个靶基因启动子内的激素应答元件。受体反式激活结构域负责在启动子上与乙酰转移酶类、共激活因子以及通用转录分子组(TBP、TFIIB、RNA 聚合酶 II)相互作用,进而导致转录激活。
II 型非类固醇核受体包括甲状腺激素受体、视黄酸受体,维生素 D 受体和 PPARγ。该家族的成员可异质二聚体化类视黄醇 X 受体 (RXR)。在结合配体前,受体异二聚体在胞核作为与组蛋白脱乙酰酶 (HDAC) 和其他共抑制因子形成的复合体的一部分,该复合体可将靶标 DNA 限制在一个紧密缠绕的构象内,以防暴露在反式激活因子下。配体结合会导致 HDAC 解离、染色质去抑制以及转录激活。
除配体结合外,多种许多生长因子和细胞因子信号转导级联的作用也可调节核受体活性,其中,信号转导级联会导致受体磷酸化或其他翻译后修饰,尤其是在氨基末端反式激活结构域内。例如,雌激素受体会在多个能够影响受体活性的丝氨酸残基上被磷酸化。Ser118 可以是转录调节激酶 CDK7 的底物,其中 Ser167 可被 p90RSK 和 Akt 磷酸化。Ser167 位点的磷酸化可能使乳腺癌患者产生它莫西芬耐药性。