您可以通过点击 CST 信号转导通路图的各个节点查找研究资源或产品信息。您还可以下载通路图,将其用于教育和研究。

细胞骨架由三种胞内纤维构成:微管、微丝(肌动蛋白微丝)和中间纤维。细胞骨架信号转导调控数个重要的细胞进程,例如:细胞分裂、黏附、极性、迁移以及通过纤毛和鞭毛的运动。
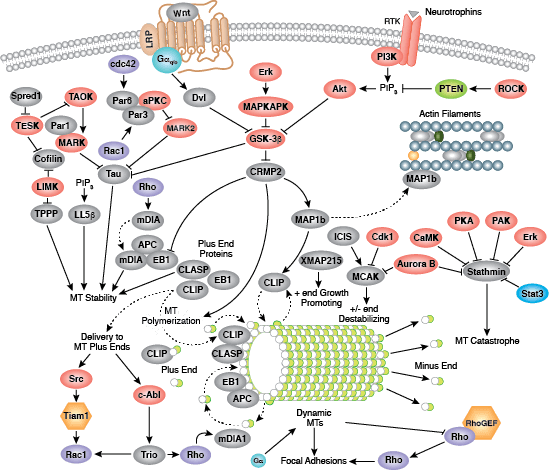
微管包含球形微管蛋白亚基,由 α/β-微管蛋白异二聚体形成所有真核细胞的微管蛋白亚基。要形成微管蛋白亚基的成核聚合,进而形成微管聚合物,γ−微管蛋白是必要的。许多细胞运动是通过微管动力介导的,包括纤毛和鞭毛的跳动、膜泡的核质转运和神经轴突的迁移。微管也在有丝分裂/减数分裂过程中的纺锤体组装中发挥关键作用,而且负责中期的染色体排布。微管组成中心粒的 9 + 2 结构,这是作为微管组织中心 (MTOC) 发挥作用并影响细胞极性的中心体的关键部分。正是由于微管在有丝分裂中的作用,微管已经成为癌症化疗中的靶标。微管持续经历动态不稳定性过程,正极的微管聚合与负极的解聚合进行竞争。这个过程由数个诸如 stathmin、diap1/2、tau 以及小 GTPase 的 Rho 家族的信号转导分子进行调控。
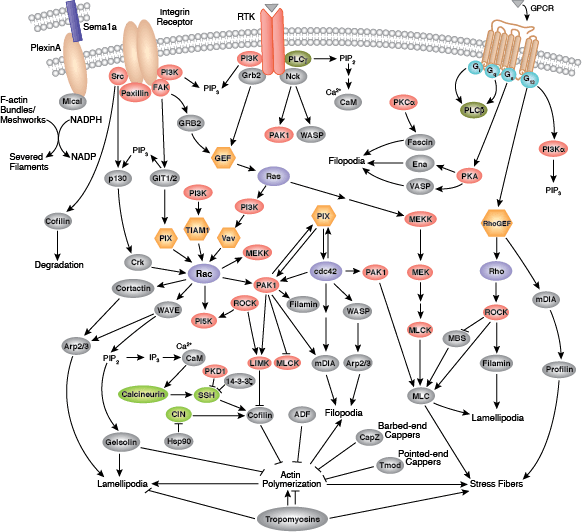
微丝是细胞骨架的主要结构部分,由肌动蛋白、丝状肌动蛋白的纤维聚合物组成。微丝对于细胞形状变化、迁移、增殖和存活至关重要。调控肌动蛋白细胞骨架始于通过 G 蛋白偶联受体 (GPCR)、整合素、受体酪氨酸激酶 (RTK) 以及许多其他诸如 semaphorin 1a 受体 PlexinA 的专门受体进行的信号转导。这些受体会启动大量信号转导级联反应,包括小 GTPase 的 Rho 家族(Rho、Rac 和 Cdc42)及其激活剂、鸟嘌呤核苷酸交换因子 (GEF)、它们的下游蛋白激酶效应器,包括 Rho-kinase/ROCK 和 p21 激活的激酶 (PAK) 以及通过 GTPase 与数个诸如皮动蛋白、diap1/2、WAVE 和 WASP 的肌动蛋白调节蛋白的直接结合。这些蛋白质上的级联聚合直接调控肌动蛋白细胞骨架的行为与组织,包括肌动蛋白相互作用的调节蛋白,例如:cofilin、Arp2/3 复合体、Ena/VASP、抑制蛋白和凝溶胶蛋白。
中间纤丝的主要类型根据其细胞特异性表达进行区分:细胞角蛋白(上皮细胞)、胶质原纤维酸性蛋白或 GFAP(胶质细胞)、肌间线蛋白(骨骼细胞、内脏细胞和某些血管平滑肌细胞)、波形蛋白(间充质来源)和神经纤维细丝(神经细胞)。该组的成员都具有一个球状的氨基端头结构域、一个中央 α−微管蛋白杆结构域和一个羧基末端尾部。中间纤丝为细胞提供结构支撑,具有细胞器和动力分子的固定点的作用,并作为提供保护免受内在压力与环境压力的应激蛋白发挥作用。中间纤丝能够通过数种机制得到调控,包括磷酸化,这能够影响它们的活性,或者与其他中间纤丝相互作用的蛋白质组装的活性。