您可以通过点击 CST 信号转导通路图的各个节点查找研究资源或产品信息。您还可以下载通路图,将其用于教育和研究。

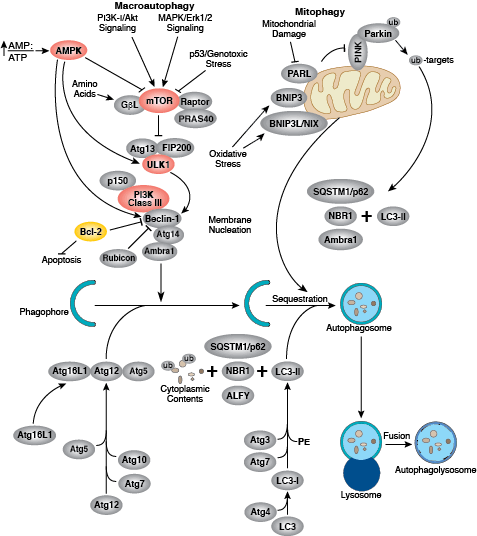
自噬是一个动态细胞循环系统,可降解细胞浆内容物、异常蛋白聚合物以及多余或受损的细胞器,以便氨基酸等基本结构可用于生成新的细胞组分。当要降解的蛋白、细胞器或细胞浆内容物被小部分膜包围时,就会出现自噬,从而产生自噬体。自噬体随后与溶酶体融合,产生一个自噬性溶酶体并导致细胞组分通过溶酶体酶被降解。通常,营养不足的条件会激活自噬,但自噬还与发育、分化、神经退行性疾病、应激、感染、肥胖及癌症等许多生理和病理过程有关。
mTOR 激酶是诱导自噬的重要调节分子,激活的 mTOR(Akt 和 MAPK 信号转导)可抑制自噬,而 mTOR(AMPK 和 p53 信号转导)的负性调节则促进自噬。已在酵母中大量发现自噬的分子机制,该机制还受大量自噬相关 (Atg) 基因的调节。丝氨酸/苏氨酸激酶 ULK 与酵母 Atg1 的作用相似,均可在 mTOR 复合体的下游发挥作用。ULK 与 Atg13 和支架蛋白 FIP200 形成一个较大的复合体。III 类磷酸肌醇 3-激酶(III 类 PI3K 或 hVps34)是自噬的一个主要调节分子。III 类 PI3K 结合多种蛋白,包括 p105/Vsp15、Beclin-1、UVRAG、Atg14 和 Rubicon。Rubicon 会抑制 III 类 PI3K 脂质激酶活性,并对抗 Atg14L 的作用,Atg14L 是 III 类 PI3K 活性的一个增强子。Atg 基因通过 Atg12-Atg5 和 LC3-II (Atg8-II) 复合体控制自噬体的形成。Atg12 通过一个需要 Atg7 和 Atg10(分别对应 E1 和 E2 样酶)参与的泛素样反应,结合到 Atg5。Atg12-Atg5 偶联物随后与 Atg16 发生非共价相互作用,从而形成一个大复合体。第二个复合体 LC3/Atg8 在其羧基末端被 Atg4 蛋白酶裂解活化,进而产生胞质 LC3-I。在一个需要 Atg7 和 Atg3(分别为 E1 和 E2 样酶)的泛素样反应中,LC3-I 偶联磷脂酰乙醇胺 (PE)。脂质化形式的 LC3,也称作 LC3-II,附着到自噬体的膜上。Sequestosome 1 (SQSTM1, p62) 是一种结合 LC3/Atg8 的泛素结合蛋白,可将含 SQSTM1 的蛋白聚合物运至自噬体进行降解,从而促进自噬。