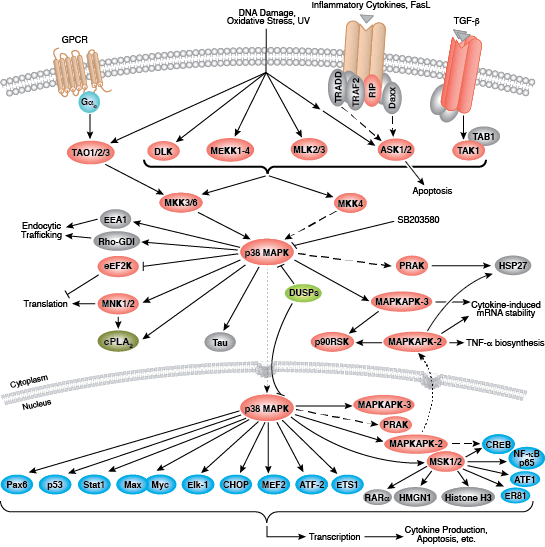
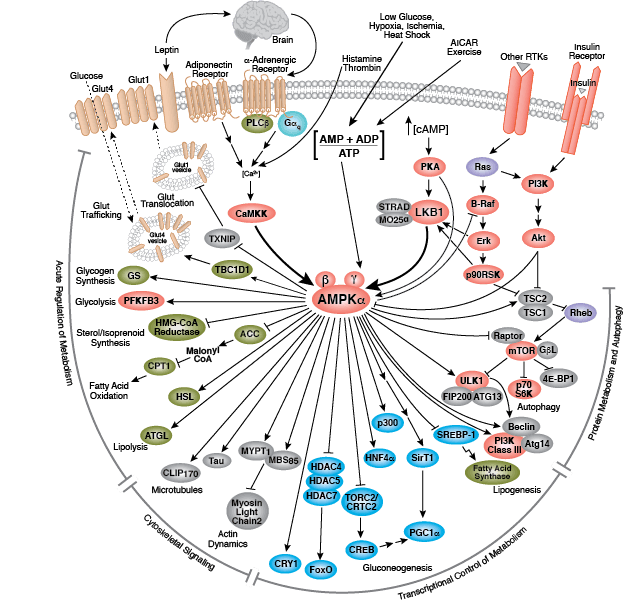
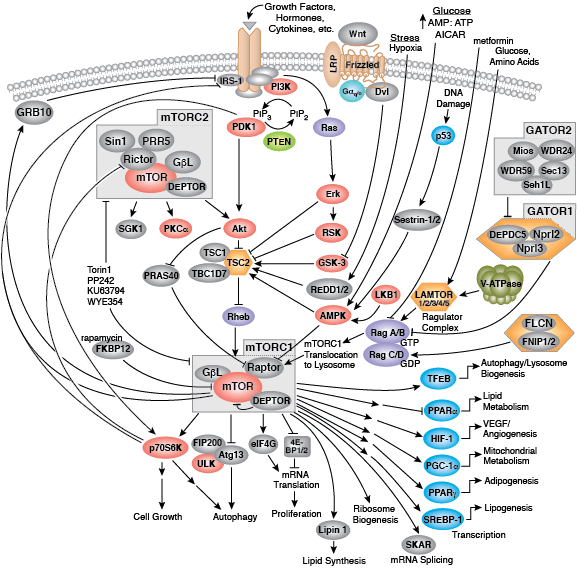
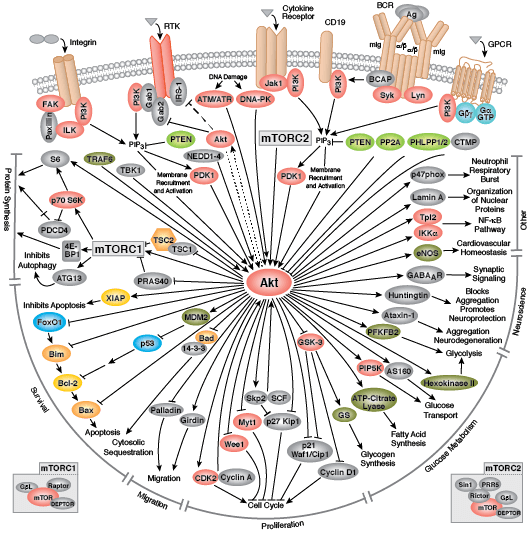
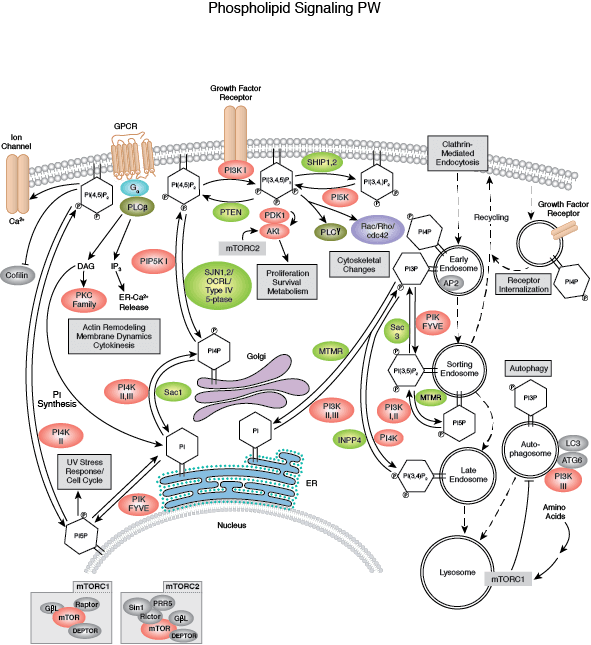
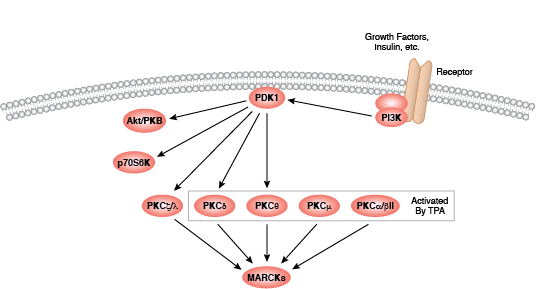
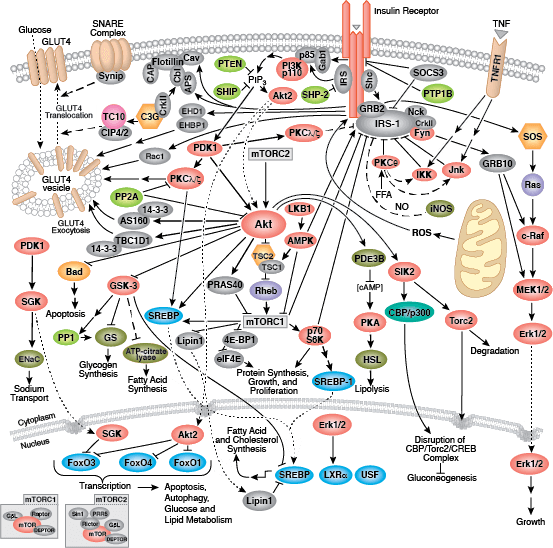
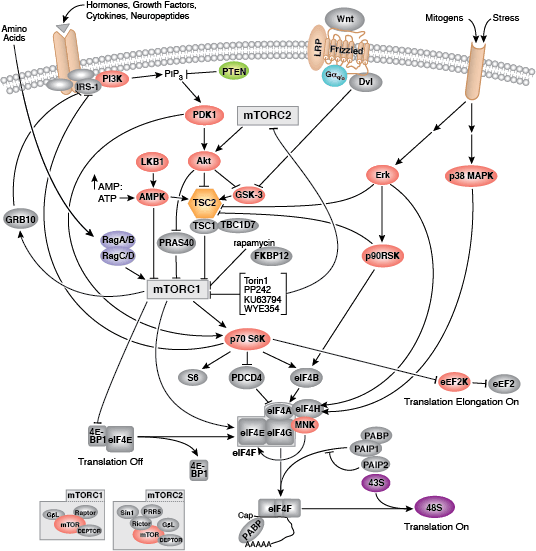
您可以通过点击 CST 信号转导通路图的各个节点查找研究资源或产品信息。您还可以下载通路图,将其用于教育和研究。

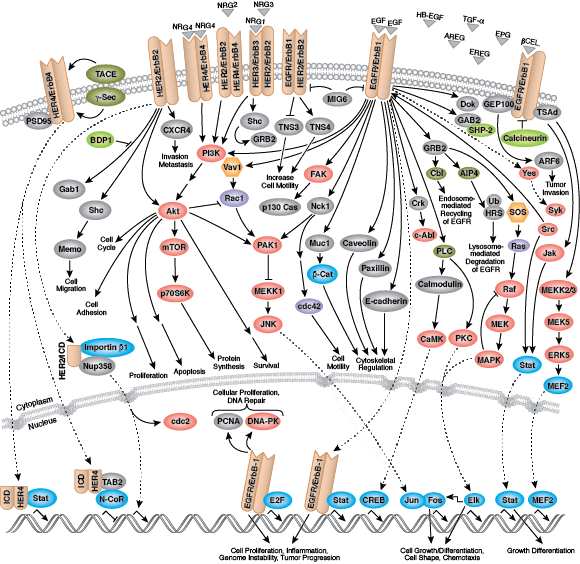
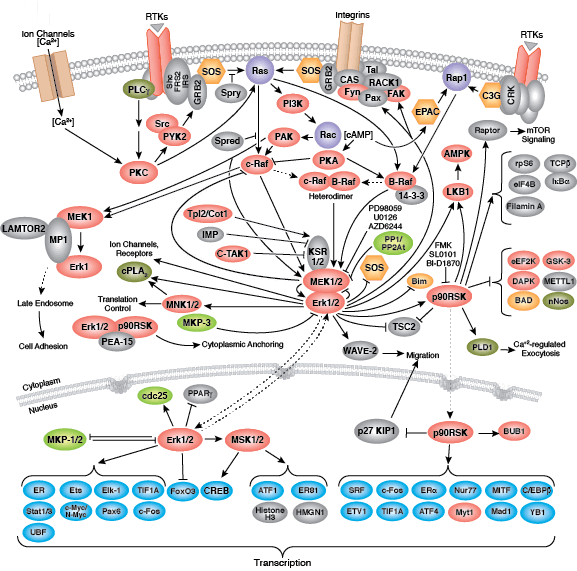
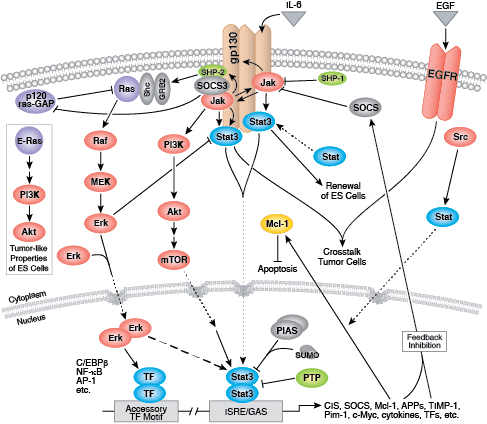
无论是作为细胞表面受体,还是作为细胞信号转导的内部效应子,酪氨酸激酶均能调控细胞和生物生长、分化和功能的多个方面。在人体中检测到约 20 个受体酪氨酸激酶 (RTK) 家族,以及至少 9 个不同的非受体酪氨酸激酶亚家族。无论定位如何,所有酪氨酸激酶可将磷酸从 ATP 转运到靶蛋白酪氨酸的羟基,从而调控靶蛋白功能。
多数受体酪氨酸激酶为单次跨膜蛋白,可结合胞外多肽配体(即生长因子)及细胞浆效应子和接头蛋白,从而调节生物进程。结合配体会促进受体二聚化和受体酪氨酸残基的自磷酸化。所导致的构象变化会使活化激酶变得稳定,并且后续的磷酸化活动会为下游接头蛋白、支架和效应蛋白形成结合位点。
表皮生长因子受体 (EGFR) 是一种广受研究的酪氨酸激酶,也是 ErbB 受体家族的一员。ErbB 蛋白结合胞外配体,并与其他家族成员形成同型二聚体或异二聚体。EGFR 和 ErbB4 结合广泛的配体,而 ErbB2 则是一个孤儿受体,没有检测到的配体。ErbB3 缺乏激活的激酶结构域,并且需要与不同 ErbB 家族成员的二聚化才能激活。结合配体后,激酶结构域活化环残基的磷酸化会维持酶活性,并为底物蛋白提供结合表面。
多种接头蛋白和效应蛋白会结合活化激酶的羧基末端尾区内的位点。在 GRB2 接头蛋白结合激活的 EGFR 磷酸化 Tyr1068 以及 Shc 支架蛋白结合磷酸化 EGFR 残基 Tyr1148 和 Tyr1173 之后,会发生 MAPK/Erk 信号转导。接头蛋白 c-Cbl 结合 EGFR Tyr1045,从而促进 EGFR 受体泛素化和降解。
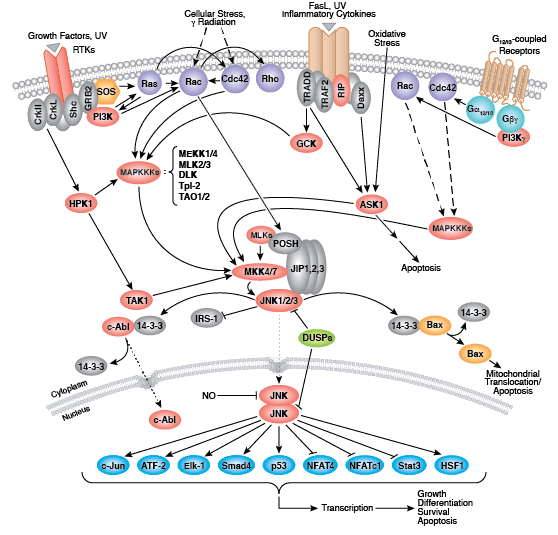
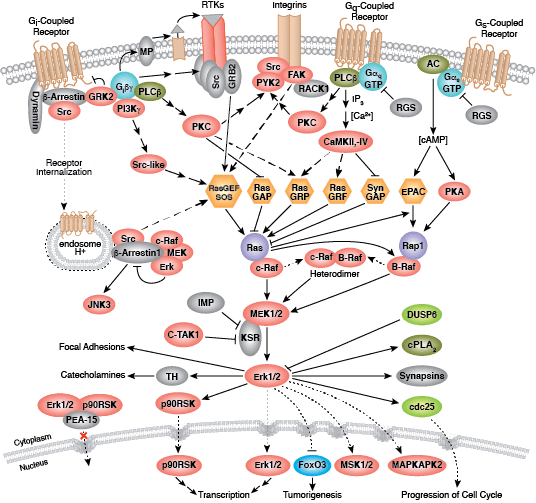
非受体酪氨酸激酶包括许多特征明显的蛋白(如 Src 家族激酶、c-Abl 和 Jak 激酶)以及其他调控真核细胞生长和分化的激酶。Tec 家族激酶(Tec、Btk、Etk、Itk 和 Txk)在 B 细胞和 T 细胞受体信号转导中非常重要,并且包含一个有助于将激酶募集到质膜的独特氨基末端普列克底物蛋白同源结构域。在天然免疫应答中,Src 家族激酶在明确激活和抑制的通路中发挥重要作用。不相关的激酶 FAK、Fer 和 ACK1 各自调节细胞运动和迁移,而 TNK1 则在负调控细胞生长的通路中发挥作用。
Zap-70 和 Syk 与免疫应答的不同方面有关,而 Fes 则在体内平衡中发挥作用。