您可以通过点击 CST 信号转导通路图的各个节点查找研究资源或产品信息。您还可以下载通路图,将其用于教育和研究。
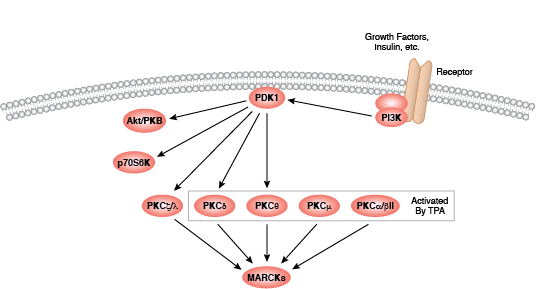
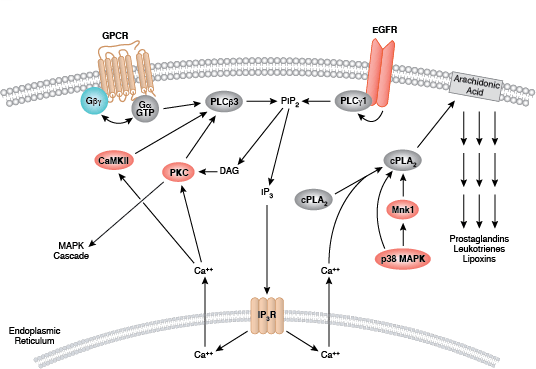
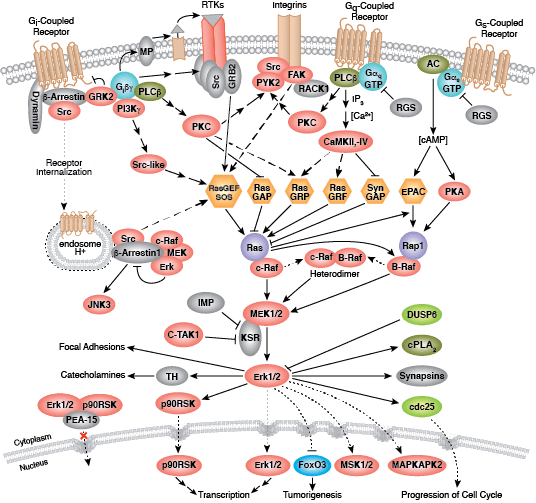
蛋白激酶 C (PKC) 家族成员可调节许多细胞响应,包括分泌、基因表达、细胞增殖和炎症反应。基本的蛋白结构包括一个氨基末端调节区,调节区通过一个铰链区连接至羧基末端激酶结构域。PKC 酶含有一个自行抑制的假性底物区域,该区域可与催化区序列结合以抑制激酶活性。第二信使激活剂的结合可将PKC 酶定位至细胞膜,细胞膜上的变构变化会使催化序列释放假底物。各种 PKC 调节区的差异使其可结合各种不同的第二信使,这也是 PKC 家族分配至 3 大组别中的依据。传统的 PKC 酶(cPKC;同工型 PKCα、PKCβ 和 PKCγ)包括功能性 C1 和 C2 调节结构域;要激活 cPKC 酶,需要将甘油二酯 (DAG) 和磷脂与 C1 结构域结合起来,同时将 Ca2+ 与 C2 结构域结合起来。新 PKC 酶(nPKC;同工型 PKCδ、PKCε、PKCη 和 PKCθ)也需要 DAG 结合以进行激活,但包含不作为 Ca2+ 传感蛋白的新 C2 结构域。最不相关的蛋白激酶 D 蛋白常与新 PKC 酶结合,因为它们会对 DAG 作出反应,但不对 Ca2+ 刺激作出反应。非典型酶(aPKC;亚型 PKCζ 和 PKCι/λ)包含非功能性 C1 结构域,但缺乏 C2 结构域,并且 aPKC 激活不需要结合第二信使。
激活 PKC 酶需要一系列磷酸化活动、与其他蛋白的结合以及变构变化。所有同工型需要在催化结构域活化环和转角基序内保守位点上磷酸化;传统酶和新型酶需要在催化结构域疏水基序内某个位点上另外磷酸化。翻译后,新生 PKC 定位至细胞膜,以结合 Hsp90 和 Cdc37。这些共伴侣分子与激酶结构域的结合先于 PDK1 对活化环位点的磷酸化作用。第二信使通过将胞质酶募集至质膜来激活 PKC,在质膜上,变构变化导致催化结构域序列释放出假底物。
其他磷酸化活动通过控制蛋白相互作用、稳定性及定位来调节 PKC 活性。例如,PKCδ 的刺激特异性磷酸化作用会导致 PKCδ 降解、剪切或活性增加。作为对 DNA 损伤和氧化应激的反应,PKCδ 通过激活 p53-介导的通路和抑制 Akt、Cdk1 和周期素 D1 等促活蛋白促进细胞凋亡。当 PKCδ 激活 Akt、NF-κB 和 MEK 通路时,转导过程通过 TNF 通路促进促活反应。