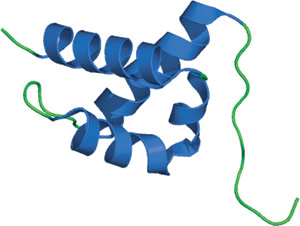
人 HYPA/FBP11 的 FF 结构域。
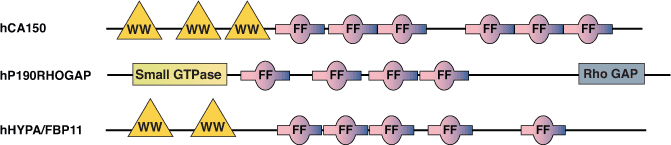
FF 结构域存在于各种各样的核转录和剪切因子以及 p190 RhoGAP 家族中。该结构域在酵母菌、人等细胞中较为保守,并且还在植物蛋白中发现。FF 结构域在氨基和羧基末端有两个保守的苯丙氨酸,结构域长约 50-60 个氨基酸,通常以串联重复序列排列。人转录因子 CA150 和剪切因子 hHYPA/FBP11 的 FF 结构域经证实会与 RNA 聚合酶 II 的丝氨酸磷酸化羧基末端结构域 (CTD) 发生相互作用。CA150 的 FF 结构域还可低亲和力地结合多种 (D/E)2/5-F/W/Y-(D/E)2/5 基序。有趣的是,所有包含 FF 结构域的胞核蛋白还在氨基末端有 WW 结构域。这可能在偶联转录和剪切的过程中发挥作用,因为这些 WW 结构域会与必要的剪切蛋白(如 mBBP/SF1)相互作用。
FF 结构域包含三个按正交束排列的 α-螺旋,其中 a 310 螺旋位于第二和第三个螺旋之间的环中。与其在串联排列中一样,氨基和羧基末端见于结构的相对末端。两个高度保守的苯丙氨酸残基在第一和第三个螺旋的中间被发现,有助于形成蛋白的疏水核心。FF 结构域的结构与其他磷酸化丝氨酸/苏氨酸结合结构域显著不同,代表着这些模块的新折叠。有人认为,螺旋 1 和 4 之间一群带正电荷的残基可介导与配体的相互作用。
| 含有 FF 结构域的 | 结合伴侣 |
| 人转录因子 CA150 | 在丝氨酸 Tat-SF1 上被磷酸化的 RNA 聚合酶 II 羧基末端结构域 (CTD) |
| 酵母剪切因子 Prp40 | 在丝氨酸上被磷酸化的 RNA 聚合酶 II 羧基末端结构域 (CTD) |
| 酵母剪切因子 HYPA/FBP11 | 在丝氨酸上被磷酸化的 RNA 聚合酶 II 羧基末端结构域 (CTD) |