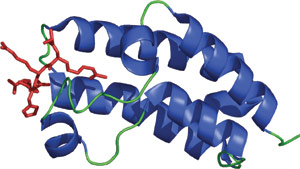
结合乙酰化赖氨酸的 Gcn5p 的溴结构域(红色)。
溴结构域在从酵母到人的一百多种蛋白中检测到,约含 110 个氨基酸,可结合乙酰化赖氨酸残基。氨基末端和羧基末端组蛋白赖氨酸尾区残基的乙酰化以及赖氨酸甲基化和丝氨酸/苏氨酸磷酸化均是重要的翻译后修饰形式,有助于在染色质组织变化期间结合组蛋白,并实现基因表达的表观遗传调控。溴结构域通常见于会调控染色质结构和基因表达的蛋白,如组蛋白乙酰转移酶以及某些核小体重构复合体的 ATP 酶组分。通过溴结构域识别乙酰化赖氨酸的模式类似于通过组蛋白乙酰转移酶乙识别酰化 CoA 的模式,但溴结构域是唯一已知会与含乙酰化赖氨酸的肽相互作用的结构域。通过溴结构域蛋白识别乙酰化赖氨酸不限于组蛋白。例如,CREB 结合蛋白转录共激活因子 (CBP) 的溴结构域可用来识别 Lys382 被乙酰化的 p53。溴结构域和乙酰化 p53 之间的相互作用发生在 DNA 损伤之前,并会促进 CDK 抑制因子 p21 的 p53 诱导性转录激活以及细胞周期停滞。
| Bromo 结构域蛋白 | 结合伴侣 | 特异性结合位点 |
| PCAF | Tat | BSYGRKAcKRRQRC |
| CBP(CREB 结合蛋白) | 三元复合体因子 Elk-1 | 未知,SSPQPKKAcKPLDGE |
| P53 | SHLKSKKAcGQSTSRHKK, SSPQPKKAcKPLDGE | |
| Gcn5p | Histone H4 | AKAcRHR |
| Celtix-1 | IRF-2 | 高乙酰化形式的 IRF-2 |