对于蛋白质表达非常低或未知的抗原靶标,可能需要在替代细胞系中使用重组蛋白或外源表达来进行抗体验证。尽管内源系统由于更能代表体内条件而被优选,但异源策略提供了许多优势。
首先,异源策略可用于验证抗体与蛋白同种型或保守家族成员的交叉反应性,从而提供有关基于抗原同源性的抗体脱靶结合潜力的有用信息。异源策略也可用于通过表达或稀释来滴定靶蛋白,从而测试抗体的敏感性。
异源策略的其他用途包括验证和优化抗体在免疫沉淀中起作用的能力,并为蛋白质印迹应用生成标准的阳性对照。此外,通过提供表达各种略有不同的蛋白质形式的选择,异源策略可以验证针对位点特异性突变体的抗体特异性。
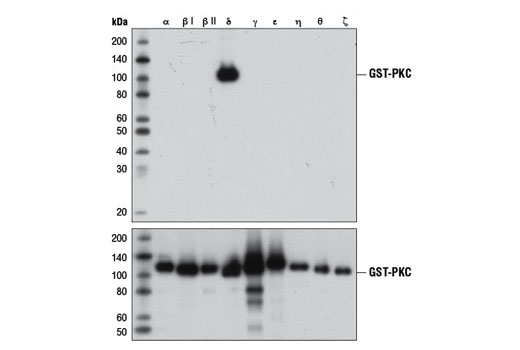
重组蛋白的普遍用途是验证抗体对一个或多个蛋白家族成员的特异性。为了说明这一点,图 1 显示了九种不同重组 PKC 同种型的蛋白质印迹分析,以确认 PKCδ (D10E2) 兔单克隆抗体的特异性。在这种情况下,抗体结合是同种型特异的,对于其他测试的重组同种型没有观察到脱靶结合。
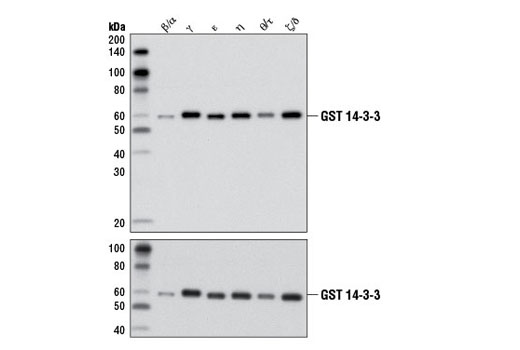
图 2 以一种稍微不同的方法证明 14-3-3(泛)抗体的泛反应性。蛋白质印迹数据清楚地显示了对所有测试的六个重组同种型的抗体识别,为同种型反应性提供了支持证据。重要的是,加入上样对照表明条带强度的差异是由于蛋白上样而不是由于抗体对不同同种型的亲和力不同所致。
这里要注意的关键点是,尽管重组蛋白或肽阵列可用于确定抗体特异性和交叉反应性,但是由于这些测定法的人工性质,这些类型的策略产生的结果可能会产生误导。此外,这些系统的一些关键功能目前尚缺乏标准化,再次强调了需要用本手册中概述的其他标志来支持任何异源策略。
设计用于在外源系统中表达目标抗原靶标的 DNA 质粒是替代使用重组蛋白的一种流行方法。同样,该策略通常用于验证抗体特异性并确定与同种型、同系物和直系同源物的交叉反应性。以这种方式表达的蛋白质易于通过已建立的技术进行分析,例如蛋白质印迹或免疫细胞化学染色。
若抗体针对表位标签、在哺乳动物细胞中未内源表达的蛋白质或仅在一组有限条件下或在稀有细胞群中表达的蛋白质,外源表达是验证此类抗体的必需步骤。由于无法在内源系统中分析这些靶标,因此外源表达对于验证抗体特异性至关重要。
图 3 显示了使用 293T 细胞制备的裂解物对 MAGE-A3 抗体进行的蛋白质印迹分析,其中 293T 细胞先采用表达各种带 Myc/DDK 标签的 MAGE 亚型载体进行转染。MAGE-A3 是一种在各种肿瘤细胞中经常表达的核蛋白,并且不是 293T 的内源性表达。这些数据说明了对 MAGE-A3 的抗体特异性,其中 DYKDDDK 抗体用于证明成功转染,而 β-肌动蛋白抗体用作上样对照。
在图 4 中,数据显示了对非天然表达蛋白 Cas9 的抗体验证的数据,其模型与预期的实验用途一致。在这里,将带有 myc 标签的 Cas9 转染到 293T 细胞中,并使用 Cas9(化脓性链球菌) (E7M1H) 兔单克隆抗体进行检测,以进行免疫细胞化学染色。使用 Myc-Tag (9B11) 小鼠单克隆抗体确认 cDNA 的表达。

因为异源策略允许表达几乎无限的构造,它不限于表达野生型蛋白。可以使用在细菌或哺乳动物细胞中外源/重组表达的蛋白质的位点特异性突变体来验证位点特异性抗体的特异性。
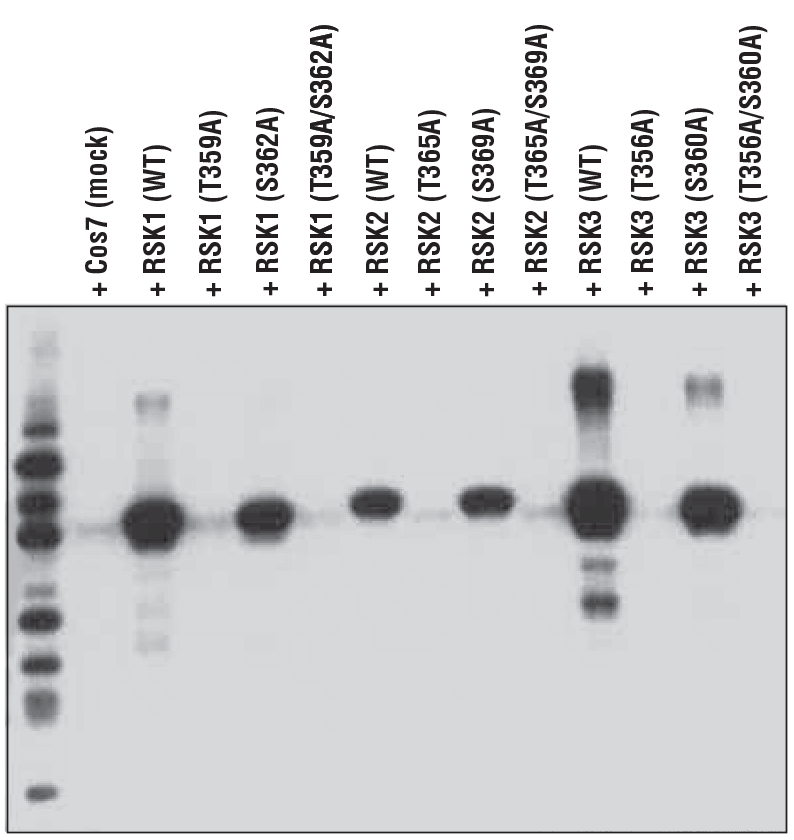
图5 说明了这一概念,证明了磷酸化 p90RSK (Thr359) (D1E9) 兔单克隆抗体对 p90RSK 蛋白的 Thr359 磷酸化形式的特异性。在这种情况下,除了 p90RSK 家族成员 RSK2 和 RSK3 上的等效位点之外,该抗体还用于探测从表达野生型蛋白、Thr359 至 Ala (T359A) 突变体、Ser362 至 Ala (S362A) 突变体或双重突变体 (T359A/S362A) 的细胞制备的裂解物。
这些结果突出了抗体的两个重要特征:苏氨酸位点的特异性和同等检测其他家庭成员保守位点的能力。这些实验很少进行或被大多数供应商忽视。
由于异源策略与其他标志的明显不同点在于它依赖于抗原靶标的非天然表达,因此将其与其他验证策略结合使用以获得关于抗体特异性和敏感性的有用认识至关重要。孤立地使用异源策略可能会产生误导,但与其他标记一起使用,它代表了一种非常强大的抗体验证方法。